Genetics 08: 'Bacterial Basics, Phages, and CRISPR elements'
These are my notes from lecture 08 of Harvard’s Genetics 201 course, delivered by Tom Bernhardt on September 24, 2014.
Tom’s reasons to care about bacteria:
- Pathogenesis.
- Basic science. Signaling, translation, evolution.
- Synthetic biology. Bioremediation, biofuels.
- Food production
Bacteria basics
The biggest division in bacteria is gram negative vs. gram positive.
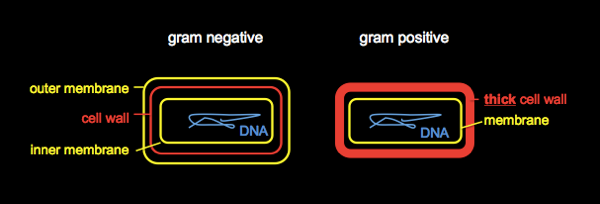
Examples of gram negative bacteria include E. coli, V. cholerae, Salmonella, Shigella. Examples of gram positive include Staph. aureus, Streptomyces, B. subtils, Lactobacillus. Streptomyces is responsible for the smell of soil.
| Property | Human | Yeast | E. coli |
|---|---|---|---|
| Genome size (bp) | 3e9 | 1.2e7 | 4e6 |
| Number of genes | ~20K | ~6K | ~4K |
| Cell diameter | 10-20 μm | 5 μm | 1 μm (and 2-5μm length) |
The bacterial genome consists of 1 circular chromosome with 1 origin of replication, haploid, usually no introns. DNA is unenveloped in the cytoplasm. Bacterial genes are often organized as follows. A single operon controls multiple genes. One promoter runs from positions -35 to -10 relative to the transcription start site of the first gene in the array. Sigma factors bind the promoter and recruit RNA polymerase.
Plasmids can be used to transfer information between bacteria via transformation or conjugation. Antibiotic resistance cassettes such as ampicillin, streptomycin, kanamycin or chloramphenicol resistance can be used to select clones of interest. Phages can be used to transduce genes and get genome integration.
Phages
The bacteriophage life cycle is as follows:
- Bind a receptor on the outside of the bacterium.
- Inject DNA into host cell.
- Express genes.
- Replicate the genome.
- Form capsids.
- Assemble whole viruses from genomes and capsids.
- Lyse the cell to get out again.
For bacteriophage λ for instance, one infected cell can produce ~100 viruses, with the whole life cycle lasting about 40 minutes (in LB at 37°C).
Fun numbers:
- Bacterial cells on earth: 1e30
- Phages on earth: 1e31
- The 1e31 phages contain DNA totaling 200 million light years in linear dimension.
- The entire earth’s bacterial population turns over every 2 days, mostly due to phage infection. This corresponds to 1e25 infection events per second.
A classic assay for studying phages is the “plaque assay”. Start with a phage stock and create serial 10-fold dilutions. Introduce each dilution into a bacterial culture, let them infect for, say, 20 minutes, then add 4mL of soft (warm and molten, but not hot enough to kill cells) agar, then put it onto a petri plate coated with LB agar. By the next morning you’ll have a full bacterial lawn on the plate if the bacteria are healthy (weak dilutions), and a totally clean plate if there were enough phage to completely destroy all of the bacteria (strong dilutions). At intermediate dilutions you will have a lawn interspersed with dots (“plaques”) which represent sites of replication of individual clones of phage. The plate which has this dotted pattern is the limiting dilution, and from the number of such plaques can be used to infer the concentration of phage in the original stock. With many phages, the plaques are clear, representing sites of no bacterial growth. Other phages integrate into the host genome and the bacteria thus stably infected become resistant to further infection, and can live, thus the plaques appear turbid.
A typical overnight E. coli culture contains 1e9 cells per mL.
Some phages can only replicate by lysis. Such phages are called virulent. Other phages can replicate either by lysis or by lysogeny. For instance, bacteriophage λ’s DNA has sticky ends which allow it to circularize after injection into the host cytoplasm, and it also has an attachment site called attP. The bacterial genome has a corresponding site called attB. A viral site-specific recombinase will cause crossover between these two sites, catalyzed by integrase (int gene in λ), integrating the viral genome into the host genome. Now the viral genome is replicated whenever the cell divides. The viral genome encodes a repressor which binds most of the viral gene promoters and silences them. The phage DNA in the host genome is called a prophage and the cell is called a lysogen. The repressor will also silence any incoming DNA from additional phages that attack the cell, thus preventing infection by additional phages. Phages capable of forming lysogens are called temperate phages. Temperate phages tend to contain a mechanism for prophage induction, where if it senses a danger signal (e.g. DNA damage in the bacterium) it will convert to lytic replication.
Site-specific recombination using λ’s proteins is the basis for gateway cloning. Many bacterial strains that are pathogenic to humans, such as E. coli 0157, are lysogens, and the sickness-causing toxins they produce are actually encoded in viral genes.
CRISPR
Phages are the biggest predators of bacteria, and so much of bacterial behavior is evolved to counter phages. Evolution against phages led to restriction enzymes. Bacteria produce restriction enzymes to cleave phage DNA, and then the bacteria methylate their own genomes, which makes them resistant to their own restriction enzymes. CRISPR is another such defense mechanism. Ishino first reported in Journal of Bacteriology the identification of some strange repeating 20-30 nt sequences in an E. coli genome [Ishino 1987]. The significance of these repeats was not clear until 2002 when it was revelead that such repeats were common to many bacterial genomes, and were found adjacent to a set of open reading frames [Jansen 2002]. Jansen dubbed the repeats Clustered Regularly Interspaced Short Palindromic Repeats (CRISPRs), and named the ORFs CRISPR-associated or “Cas” genes. In between the repeats were sequences from phage plasmids. It was eventually hypothesized that this represented an immune system targeting phage DNA, and an analogy to RNAi was later suggested [Makarova 2006].
A promoter upstream of the CRISPR sequences induces transcription of a CRISPR RNA which forms stem loops. The stem loops are processed into small RNAs which are bound by Cas protein. The small RNAs bind phage DNA, and Cas is a nuclease which cuts the phage DNA. The target sequence in the phage has to have an adjacent motif called a PAM (protospacer adjacent motif) in order to be cut. The PAM is not present in the copy of the DNA stored in the bacterial genome, which is how the bacteria avoid cutting their own DNA.
CRISPR is now in use to edit genomes by cutting with Cas’s nuclease activity, or to silence genes by simply binding but not cutting DNA using a defective Cas called dCas9 - this technique is called CRISPRi (CRISPR interference).
An example genetics experiment
At the end of class we started an example genetics experiment to test the hypothesis that the CRISPR system confers immunity to phages. Please see the lecture 09 notes.

About Eric Vallabh Minikel
Eric Vallabh Minikel is on a lifelong quest to prevent prion disease. He is a scientist based at the Broad Institute of MIT and Harvard.