PrP antibodies in humans: the experiments of nature
introduction
A powerful thing about human genetics is that it allows us to read out the experiments of nature that have already been done. If you want to know what happens if you tinker with this gene or that, sequencing enough people can sometimes give you an answer. Genetics can thus be a lever to understanding the true mechanism of disease or designing a drug that works. But a great many of nature’s genetic experiments come in the form of expression or activity — meaning, genetic variants that increase or decrease a gene’s expression level or a protein’s activity. This might predict what would happen if you raise or lower the activity of that gene’s product, but it may not provide as clear of insight into what would happen if you bind that gene’s product with a therapeutic ligand. At Prion2019, Adriano Aguzzi articulated a vision: that by studying naturally occurring antibodies in the human population, we can learn the effects of binding a particular target, and gain some of those insights that genetics might miss. Through a project at University Hospital Zurich and his spinout company Mabylon, he is doing high-throughput automated screening of blood samples over 100,000 people per year to figure out which proteins they have circulating antibodies against, and what clinical outcomes those antibody profiles might correlate with.
I was fascinated by this idea, and have been following with great curiosity the recent findings from his and other labs to see what we can learn about my target of interest, PrP. Sonia’s and my work at the Broad is focused primarily on lowering PrP, but another therapeutic hypothesis in prion disease is to bind PrP with a small molecule or an antibody. In principle, binding PrP in its normal conformation should be able to stabilize it against misfolding, and prevent prion propagation. The idea is elegant, and it’s been validated in cell culture: monoclonal antibodies (mAbs) against PrP clear prion infectivity in cells [Enari 2001, Peretz 2001], and “stapling” together the subdomains of PrP via non-native disulfide bonds (which might simulate the effect of a binder) can also confer resistance to prion infection [Hafner-Bratkovic 2011]. But there remain unanswered questions about this strategy. First, can it work in vivo? In animals peripherally infected with prions, PrP mAbs can be curative if administered early enough [White 2003], but unfortunately, mAbs don’t seem to work once prions are in the brain. Whether that’s due to pharmacokinetics (the difficulty of getting enough antibody distributed widely enough across the brain) or some more fundamental biological reason, we don’t yet know. Second, what epitopes (what parts of PrP’s surface) can be safely targeted? mAbs to certain epitopes of PrP’s C-terminal globular domain can be toxic at high doses, and while it’s not yet clear whether those toxic doses are relevant for therapy, it’s been shown pretty clearly that the toxicity is on-target (mediated by PrP binding) which might be mechanistically interesting [Sonati 2013, Reimann & Sonati 2016]. As of last year, the PRN100 antibody has been used clinically in six patients, and although it was not a clinical trial and not many data are available yet, any results are likely to influence the future development of PrP mAbs. Against this backdrop, any data on naturally occurring antibodies to PrP in humans could be grealty informative.
Of course, Aguzzi is far from the first to look for naturally occurring antibodies in humans. In fact, blood donated for transfusion is routinely screened for “blood group”, which, I am learning, encompasses far more than just A/B/O and Rh +/-. The International Society of Blood Transfusion currently recognizes 35 blood group systems based on red cells alone. What this means is that even if two people are ABO and Rh-matched, one peron might still have antibodies against some other antigen that the other person presents, such as Duffy, and this can result in transfusion incompatibility. Some people are said to have “rare blood”, which can be an enormous challenge if you also have a bleeding disorder and require a lot of transfusions. If you receive incompatible blood, your immune system attacks it and destroys it (hemolysis), and what’s worse, it may over time develop new antibodies against other antigens found on that blood, further narrowing the set of possible blood donors you could receive from in the future. A related problem is that sometimes a mother’s antibodies can attack the blood of her baby, resulting in hemolytic disease of the newborn.
PrP antibodies in E219K homozygotes
Beginning in 1991, researchers in Japan took note of a never-before-seen blood group they dubbed “anti-KANNO” [Kawabata 2014]. The antigen (KANNO) was high-frequency, meaning, it is found in almost everyone’s blood, while the antibody (anti-KANNO) was super rare, with just 28 cases identified in all of Japan and none reported elsewhere. All of the people with the anti-KANNO blood group were women who had been pregnant, and although only five of their babies were available for testing, those babies were all negative for anti-KANNO. This could suggest the possibility that the women had developed antibodies against an antigen present in their babies during pregnancy. Luckily there did not seem to be any major medical consequence to the anti-KANNO blood group. Several women underwent transfusions, and although their blood tested as being reactive to the transfused blood, it did not result in hemolysis. Similarly, none of the babies delivered seemed to be suffering hemolytic disease of the newborn. So anti-KANNO was a documented molecular phenomenon, but not a disease and not even really a phenotype per se.
Where it gets interesting is that last year, a group of researchers did an exome sequencing and genotyping study to search for a genetic basis of anti-KANNO [Omae & Ito 2019]. This is the kind of genetic study that could easily turn up nothing: they analyzed DNA from just 22 anti-KANNO cases and a few hundred controls, so there was the risk that with limited statistical power, they might only be able to see a smoking gun. But a smoking gun is exactly what they found: 22/22 cases were homozygous for the PRNP E219K polymorphism. This genetic variant, first identified as a protective factor against prion disease, has an allele frequency of just 6% in Japan [Shibuya 1998], so finding 22/22 anti-KANNO individuals homozygous is incredibly unlikely by chance, and none of the individuals positive for the KANNO antigen were homozygotes. Overall they had an estimated odds ratio of 1060, with P value of 3 × 10-8. By Western blotting, the researchers were able to show that anti-KANNO serum recognized 219E PrP but not 219K PrP, conclusively confirming that the KANNO antigen was none other than wild-type PrP [Omae & Ito 2019].
PrP residue 219 is on the surface of the C-terminal globular domain, and given that anti-KANNO is non-reactive against 219K PrP, the simplest explanation would be that the anti-KANNO antibodies bind this epitope. If true, that’s interesting because residue 219 (corresponding to residue 220 in mouse PrP numbering) lies smack in the middle of the epitope for antibodies POM4, POM10, and POM19 (the epitope for each was reported to include residues 218-221 in mouse), all of which were reported to exhibit on-target toxicity [Sonati 2013]. In the figure below, you can see this residue (yellow) in the middle of a black (toxic) epitope on PrP’s surface.
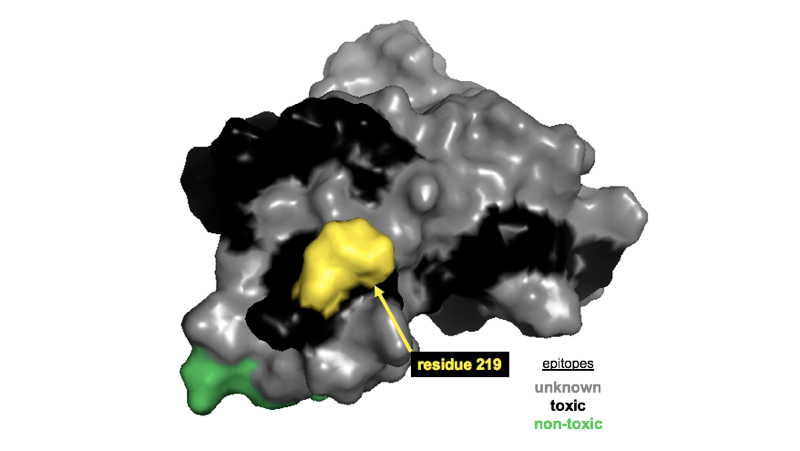
Location of residue 219 (mouse residue 220) in the context of reportedly toxic and non-toxic epitopes [Sonati 2013]. Adapted from this post.
Still, this in and of itself doesn’t tell us much about which epitopes of PrP are okay to try to bind with a drug. For one, though it seems likely that anti-KANNO antibodies bind PrP at this epitope we don’t know that for sure — it’s possible that the E219K variant causes some conformational change in PrP and that anti-KANNO binds a neo-epitope exposed by this change. More importantly, though, anti-KANNO is an alloantibody — an antibody that people have against foreign antigens, not against their own body’s antigens. So this doesn’t yet tell us anything about what would happen if people had antibodies that could bind their own PrP on their own cells. To find out what happens when people’s antibodies bind their own PrP, one wants to look at autoantibodies — those that do bind one’s own proteins.
PrP antibodies in PRNP mutation carriers and controls
A question that everyone always asks, and that no one has the answer to, is why genetic prion disease strikes when it does. How can people carry the mutation their whole lives, and yet disease strikes so suddenly later in life? Why does it come so much earlier for some than others? To date, we haven’t found any factors that predict age of onset [Minikel 2019]. And unlike Huntington’s or Alzheimer’s, where it appears the disease process is underway for decades and just takes a long time to manifest in the form of symptoms [Bateman 2012, Byrne 2018], prion diease seems to strike pretty suddenly: most pre-symptomatic mutation carriers are normal in terms of fluid biomarkers, with no molecular evidence of ongoing pathology in their blood or spinal fluid [Vallabh 2019].
A few years ago, Karl Frontzek and Adriano Aguzzi had a hypothesis: could naturally occurring autoantibodies against PrP be keeping prion disease at bay in some subset of individuals? Perhaps PRNP mutation carriers have anti-PrP antibodies that slow or prevent disease development, but the titers drop with age, triggering onset of disease. Or, perhaps some people have anti-PrP antibodies and some don’t, and the people with them are the people who develop disease later in life. To test these hypotheses, they set out to collect blood samples from people (both pre-symptomatic and symptomatic) with PRNP mutations, and mutation-negative controls from genetic prion disease families. Participants and researchers from across the world sent in blood samples (including us — we shipped samples from our MGH clinical research study). All told, the researchers were able to test blood from 46 symptomatic genetic prion disease patients, 76 pre-symptomatic mutation carriers, and 78 mutation-negative controls.
The results of this study, posted to medRxiv last year, were just officially published [Frontzek 2020]. Intriguingly, they found evidence of anti-PrP antibodies in a significant fraction of study participants — our first evidence that anti-PrP autoantibodies (as opposed to alloantibodies like anti-KANNO) do indeed occur naturally in humans. Exactly what proportion of people had anti-PrP antibodies depends on where you draw the cutoff (peruse figures e-1 and e-2 in the supplement). A few patients exhibited pretty strong reactivity (-logEC50 > 2.5), on par with the positive control of spiked-in POM1 antibody, while nearly all participants were above the level of negative controls (IgG-depleted serum; -logEC50 > 1.0). The analyses in the paper treat the measured anti-PrP titer as a continuous variable rather than using any specific cutoff.
The main finding of the study is that there is no evidence for the original hypothesis [Frontzek 2020]. Anti-PrP autoantibodies do exist, and are reasonably prevalent, but they don’t seem to correlate with any of the variables of interest. There was just a slight correlation with age, with suggestively higher titers in younger people. But antibody titers weren’t higher in symptomatic patients than in pre-symptomatic mutation carriers, nor were they higher in pre-symptomatic mutation carriers than in mutation-negative controls. This argues against the idea that these antibodies were staving off disease, or that they developed in response to PrP mutations or PrP misfolding. There also were no differences in antibody titers between people with the most common PRNP mutations, and the titers were no higher in people with autoimmune disease (of whom there happened to be 8 in the cohort) than in those without. On average, titers didn’t change much over time in the people who gave blood to the study more than once, and the rate of change was no different in mutation-positive vs. negative individuals.
Does the lack of any correlation for all the variables mentioned above provide evidence that anti-PrP antibodies aren’t a good therapeutic hypothesis in prion disease? No — the results, while interesting, stop short of ruling out this therapeutic approach. For one, the assay they used to screen people’s blood looked for reactivity against wild-type, properly folded recombinant PrP. The findings here don’t rule out the possibility that some people might have antibodies against the mutant forms of PrP, or antibodies specific to misfolded PrP. The latter possibility would be the most fascinating of all, since to date no one has been able to create in animals an antibody specific to misfolded PrP. Equally important, we still don’t know much about the relationship between anti-PrP antibodies circulating in the periphery, and any activity they might or might not have in the brain (see discussion at bottom). It’s conceivable, for instance, that some of the antibodies of people in this study do in fact possess profound anti-prion properties, but that they simply don’t enter the brain in sufficient quantity to impact the development of prion disease.
PrP antibodies in the general population
An even more recent study just posted to bioRxiv [Senatore 2020] explores the prevalence of anti-PrP autoantibodies in the general population, using that cohort mentioned at top: people who had blood drawn, for any reason, at University Hospital Zurich, and agreed to donate a sample to the study. Recall that in the smaller PRNP mutation carrier and control cohort the researchers were able to find a few people with fairly high anti-PrP antibody titers (say, 2.5 -logEC50 units) [Frontzek 2020]. Here, by casting a much wider net, they were able to find more people above this level, including a few with really high titers. They called 21 people out of 37,894 as being above 2.0 -logEC50 units, and a few were as high as 3.0 or 3.5. Just as in the PRNP mutation carrier study, there seemed to be no correlaton with clinical parameters: the people with anti-PrP antibodies had all different reasons for having been at the hospital, and were not enriched for any particular phenotype or age group. The researchers tested the reactivity of these antibodies for three different recombinant PrP constructs — full-length, N terminus only, and C terminus only — and found that some patients had antibodies only to the C-terminus while others had antibodies against both halves of the protein. This seems to suggest a polyclonal response, meaning some of these people had more than one different clonal B cell population producing more than one different antibody against PrP. The authors did not yet determine the exact epitope of each antibody, so we don’t have any resolution on whether the antibodies to the C terminus were binding the epitopes that have been deemed toxic or non-toxic — but we can conclude there are at least a couple of different places that peripheral antibodies can bind PrP and still be tolerated.
This study also employed a few other approaches to understanding how anti-PrP antibodies behave and exist in the population. Up to now, most of the studies of PrP antibody toxicity had been done using the POM monoclonals, a series of antibodies created by immunizing PrP knockout mice [Polymenidou 2008]. Here, in order to more exhaustively ascertain the set of possible antibodies to possible PrP epitopes, they used phage display libraries to pan for anti-PrP antibody fragments (Fabs) [Senatore 2020], and managed to identify some Fabs even against hard-to-target regions of PrP, such as the very tip of the N terminus. They used these to validate many of the earlier findings about which epitopes of PrP are therapeutic or toxic to bind with an antibody. They also compared the sequences of these Fabs to published sequences of human B cell repertoires from the general population, and found that some B cells encoded antibodies with a complementarity-determining region (CDR) that differed from a PrP Fab by only 1 or 2 or 3 residues. One interpretation is that those B cells were also targeting PrP. But, when those B cell sequences were cloned and expressed and the corresponding Fabs purified, they only bound PrP with micromolar affinity [Senatore 2020]. Micromolar is not super strong for an antibody — many of the the phage display Fabs bound PrP with mid- to low-nanomolar affinity, and some of the POMs have sub-nanomolar affinity [Polymenidou 2008]. The study notes that these same antibodies might have actually very strong affinity in bivalent form, which is possible, though it is also hard to rule out the possibility that the CDRs identified in those B cell repertoires were actually targeting some protein other than PrP.
conclusions
What can we learn about the therapeutic — or toxic — potential of PrP antibodies from screening for them in the general population? My take is that so far, we have learned at least two important lessons. First, naturally occurring anti-PrP antibodies do exist in the population, both as alloantibodies in E219K homozygotes and also as autoantibodies in PRNP mutation carriers and in unselected healthy controls. This rules out a most extreme scenario, in which any and all anti-PrP antibodies are super toxic even at low titers. Second, the titer of anti-PrP autoantibodies seems uncorrelated with any clinical parameters in PRNP mutation carriers or controls. This argues against a model in which the existence or degree of immune response against PrP is a principal mechanism determining age of onset in genetic prion disease.
While I am reasonably confident in those two takeaways, there remain a lot of open questions.
A major gap in my understanding — and perhaps in anyone’s understanding, though maybe I just haven’t read enough of the literature — is whether peripheral autoantibody titers predict any, and if so, how much, antibody titer in the brain parenchyma. The relationship between peripheral and brain antibody concentration has been studied in the case of passive immunization, where antibodies are infused into the bloodstream, as for a drug regimen. Some oft-cited animal studies have found that 0.02% to 0.11% of the total antibody dose ever enters the brain [Banks 2002, Levites 2006], while recent data from preclinical studies of aducanumab (Biogen’s anti-amyloid antibody in development for Alzheimer’s) show a brain-to-plasma concentration ratio of 1.3% [Sevigny 2016]. There’s a lot of variability there, and I am not sure how much is different measurements, different methods, or genuinely different properties of different antibodies. Regardless, the proportion of peripherally administered antibody that enters the brain definitely appears to be low. And although people often point to the target engagement data for aducanumab as evidence that antibodies can work as drugs for brain diseases [Sevigny 2016], it is hard to generalize that lesson across proteins (such as Aβ versus PrP) that have different localization, different expression levels, and different epitope reactivity (e.g. aducanumab is specific to misfolded Aβ whereas all known PrP antibodies bind PrPC). The fact remains that no one has yet been able to show that a peripherally administered antibody can be effective against prion disease in an animal model. Therefore, we do not yet know how what concentration of antibody would have to reach the brain in order to meaningfully treat prion disease, nor indeed, whether there even exists such a threshold. Moreover, I don’t know — maybe an immunologist would? — what concentration (in terms of molarity or mass per volume) a “high titer” autoantibody corresponds to. Thus, even if brain concentrations are 1.3% of peripheral concentrations, does the stoichiometry work out such that a “high titer” individual has appreciable antibody activity in the brain? If not, then the lack of correlation between anti-PrP autoantibody titers and clinical outcomes doesn’t rule out the possibility that those same antibodies could be beneficial — or, for that matter, harmful — if they were ever administered to the brain in large doses.
Above, I’ve been taking the newly discovered existence of these autoantibodies in the population as a given, and discussing to what degree these data provide proof-of-concept for whether an antibody against PrP could be beneficial, or toxic. But to flip it around, the other question one can ask is why some people have anti-PrP autoantibodies — what triggered their immune systems to recognize PrP in the first place? Here, too, I am not sure that we can answer the question based on any currently available data. The most recent paper [Senatore 2020] speculates that “it is not PrPC that induces adaptive immune responses against prions, but nascent PrPSc instead”, and that this “suggests a link to the low incidence of spontaneous prion diseases in human populations”. If I read and understand this correctly, the conjecture here is that PrP began to misfold in these individuals (the 21 high-titer people identified in the general population sample of 37,894), and that the immune system reacted with antibodies to PrPSc, halting the misfolding cascade and preventing them from developing sporadic CJD. Because the disease was ultimately prevented, the presence of the antibodies is not correlated with prion disease. The immunoreactivity against PrPC observed in this study would then be merely a side effect, collateral damage of the immune system’s successful war against PrPSc. Implicit is the assumption that the immune system ever even gets to see nascent PrPSc. To me, although we may never know for sure, the available data suggest that sporadic prion disease starts in the brain, not in the periphery. That said, aducanumab was discovered as a naturally occurring human autoantibody against Aβ, whose misfolding also presumably begins in the brain, so it appears not impossible for peripheral B cells to react to protein misfolding in the brain. Still, another question is, if the immune system is capable of mounting responses against nascent PrPSc, then shouldn’t we see evidence of this in PRNP mutation carriers? If the immune system really fights off prion infection by producing anti-PrP antibodies in 0.06% of people in the general population who might otherwise develop sporadic CJD, shouldn’t it also be attempting this feat in PRNP mutation carriers at 90% lifetime risk? Surely we would see some elevated rate of anti-PrP autoantibodies among mutation carriers, or some correlation with symptom onset. In any event, the hypothesis that anti-PrP autoantibodies are left over from an immune response to PrPSc is interesting, but it seems hard to prove or disprove, and unnecessary to explain the observed data.
As for the core question of whether injection of anti-PrP monoclonal antibodies could be a safe and effective therapeutic approach for prion disease, the biggest obstacle remains the lack of in vivo proof of concept for their efficacy. Without knowing what (if any) is the therapeutic concentration of anti-PrP mAb in the brain, and how (if at all) we can achieve it therapeutically, we cannot really answer the question of what epitopes they would need to target, whether there exists a therapeutic index even for toxic epitopes, how early and how often the mAbs would need to be dosed, and so on. All that being said, I do think that these “experiments of nature” have already given us some interesting food for thought, and I’m very curious to see what else we can learn from these studies over the coming years. Finer mapping of epitopes and reactivities of these naturally occurring antibodies could reveal interesting properties, and new correlations could emerge as the number of individuals studied grows. I, for one, could not have predicted years ago many of the surprising lessons that human genetic “experiments of nature” have now taught us, and perhaps antibody-based “experiments of nature” will have surprises in store for us too.

About Eric Vallabh Minikel
Eric Vallabh Minikel is on a lifelong quest to prevent prion disease. He is a scientist based at the Broad Institute of MIT and Harvard.